关于 DNA 结构发现的三篇核心文章
这三篇关于 DNA 结构的划时代文章全部发表在同一期《自然》杂志上。具体信息如下:
- 期刊期号:Nature, No. 4356(第 171 卷)
- 出版日期:1953 年 4 月 25 日
第一篇:理论模型
核酸的分子结构:脱氧核糖核酸的一种结构(第 737–738 页)
作者: J. D. Watson & F. H. C. Crick
注:此篇为 DNA 双螺旋模型的首次提出
我们希望为脱氧核糖核酸(D.N.A.)的盐提出一种结构 。这种结构具有新颖的特征,具备相当大的生物学意义 。
此前,鲍林(Pauling)和科里(Corey)已经提出了一种核酸结构 。他们慷慨地在出版前向我们提供了其手稿副本 。他们的模型由三条相互缠绕的链组成,磷酸基团位于纤维轴附近,而碱基位于外侧 。在我们看来,这种结构并不令人满意,原因有二:
- 我们认为给出 X 射线图谱的物质是盐,而非游离酸 。如果没有酸性氢原子,目前尚不清楚是什么力量将结构维持在一起,尤其是靠近轴线的带负电的磷酸基团会相互排斥 。
- 部分范德华距离似乎过小 。
弗雷泽也提出了另一种三链结构 。在他的模型中,磷酸基团位于外侧,碱基位于内侧,通过氢键连接在一起 。正如所述,该结构定义较为模糊,因此我们不对其进行评论 。
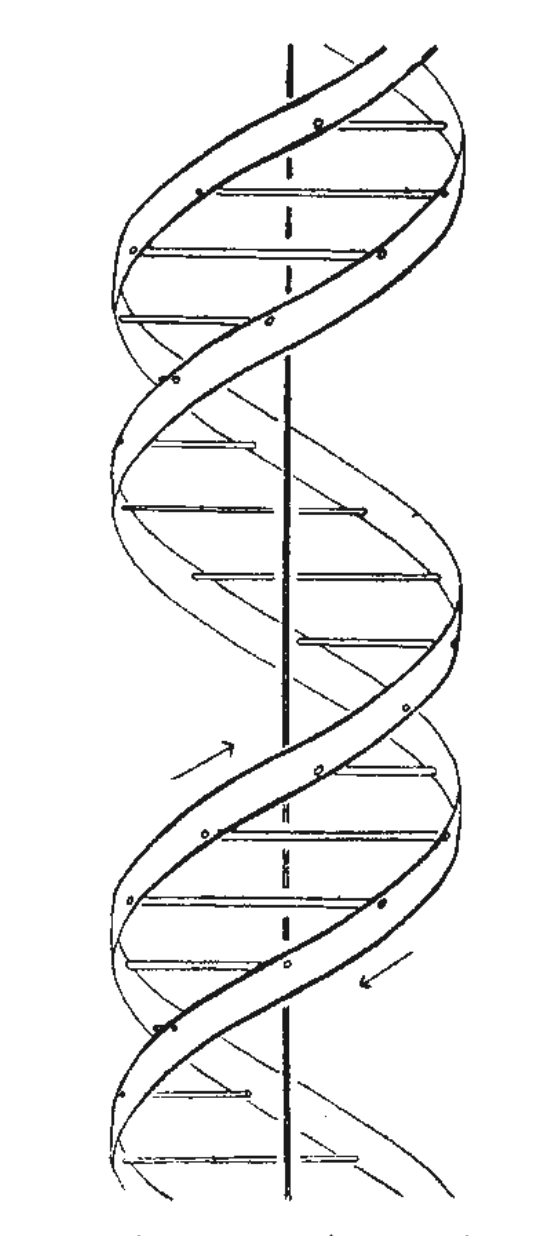
本图仅为示意图。两条丝带象征着两条磷酸-糖链,水平横杆则代表将这些链结合在一起的碱基对。垂直线标示了纤维轴。
我们希望为脱氧核糖核酸盐提出一种截然不同的结构。该结构具有两条绕同一轴线盘旋的螺旋链。我们做出了常规的化学假设,即每条链都由磷酸二酯基团组成,连接着具有 3',5' 键合的 β-D-脱氧核糖残基。
这两条链(但不包括其碱基)通过垂直于纤维轴的二回轴相关联。两条链都遵循右旋螺旋,但由于二回轴的存在,两条链中原子的序列方向相反。
每条链大致类似于富伯格(Furberg)的 1 号模型;也就是说,碱基位于螺旋内侧,而磷酸基团位于外侧。糖基及其附近原子的构型接近富伯格的“标准构型”,糖平面大致垂直于相连的碱基。
在 $z$ 轴方向上,每条链每隔 3.4 Å 就有一个残基 。我们假设同一条链上相邻残基之间的夹角为 36°,因此结构每 10 个残基重复一次,即经过 34 Å 后重复 。磷原子距纤维轴的距离为 10 Å 。由于磷酸基团位于外侧,阳离子很容易接近它们 。
该结构是开放式的,含水量相当高 。在含水量较低时,我们预期碱基会发生倾斜,从而使结构变得更加紧凑 。
该结构的新颖之处在于两条链通过嘌呤和嘧啶碱基结合在一起的方式 。碱基平面垂直于纤维轴 。它们成对结合,来自一条链的一个碱基与来自另一条链的一个碱基通过氢键相连,使得两者并排排列,且具有相同的 $z$ 坐标 。为了实现键合,这对碱基中必须一个是嘌呤,另一个是嘧啶 。氢键的形成方式如下:嘌呤 1 位对嘧啶 1 位;嘌呤 6 位对嘧啶 6 位 。
如果假设碱基仅以最合理的互变异构形式(即具有酮式而非烯醇式构型)存在于结构中,那么可以发现只有特定的碱基对才能结合在一起 。这些配对是:腺嘌呤(嘌呤)与胸腺嘧啶(嘧啶),以及鸟嘌呤(嘌呤)与胞嘧啶(嘧啶) 。换句话说,如果在任何一条链上有一个腺嘌呤成对,那么根据这些假设,另一个成员必须是胸腺嘧啶;鸟嘌呤和胞嘧啶同理 。单链上的碱基序列似乎不受任何限制 。然而,如果只能形成特定的碱基对,那么只要给定一条链上的碱基序列,另一条链上的序列就被自动确定了 。
实验发现,脱氧核糖核酸中腺嘌呤与胸腺嘧啶含量的比值,以及鸟嘌呤与胞嘧啶含量的比值,总是非常接近于 1 。
在该结构中用核糖代替脱氧核糖可能是不可能的,因为额外的氧原子会导致范德华接触过近 。
此前发表的脱氧核糖核酸 X 射线数据不足以对我们的结构进行严格检验 。就目前我们所知,它与实验数据大致相符,但在通过更精确的结果检验之前,必须将其视为未经证实的 。其中部分结果见后续通讯 。我们在设计该结构时并不了解那里呈现的结果细节;我们的结构主要(虽然不完全是)基于已发表的实验数据和立体化学论证 。
我们注意到,我们所假设的特定配对方式,立即暗示了遗传物质一种可能的复制机制。
结构的完整细节,包括构建它时所假设的条件,以及一套原子的坐标,将另行发表 。
J. D. 沃森
F. H. C. 克里克
剑桥大学卡文迪许实验室,医学研究委员会生物系统分子结构研究小组
4月2日
第二篇:普遍性证据
脱氧戊糖核酸的分子结构(第 738–740 页)
作者:M. H. F. Wilkins, A. R. Stokes & H. R. Wilson
虽然脱氧戊糖核酸(DNA)的生物学特性暗示其分子结构极其复杂,但本文描述的 X 射线衍射研究表明,其基本分子构型其实非常简单 。本通讯的目的在于初步描述一些实验证据,证明多核苷酸链的构型是螺旋状的,并且在自然状态下也以这种形式存在 。更详细的工作报告将于近期发表 。
脱氧戊糖核酸的结构在所有物种中都是相同的(尽管氮基比例差异很大),无论是在核蛋白中、提取出来的、在细胞内的,还是在纯化的核酸盐中 。相同的一组线性多核苷酸链可以以不同的方式平行堆积,形成晶体、半晶体或准晶(paracrystalline)材料 。在所有情况下,X 射线衍射照片都由两个区域组成:一个主要由沿链的核苷酸规则间距决定,另一个由链构型的较长间距决定 。沿链的不同氮基序列是不可见的 。
定向的准晶脱氧戊糖核酸(即下一篇富兰克林与葛斯林论文中的“结构 B”)给出的纤维图谱如图 1 所示 。阿斯特伯里(Astbury)曾认为强烈的 3.4 Å 反射对应于沿纤维轴的核苷酸间重复 。然而,34 Å 的层线并非由于多核苷酸组成的重复,而是由于链构型的重复引起的,因为核苷酸链的密度高于间隙水,从而产生了强烈的衍射 。子午线上或其附近反射的缺失立即提示了一个轴线平行于纤维长度的螺旋结构 。
可以证明(斯托克斯,未发表),沿螺旋等间距排列的一系列点的衍射图谱强度分布由贝塞尔函数(Bessel functions)的平方给出 。一个均匀的连续螺旋会产生一系列层线,其间距对应于螺旋螺距,第 $n$ 条层线上的强度分布与 $n$ 阶贝塞尔函数 $J_n$ 的平方成正比 。
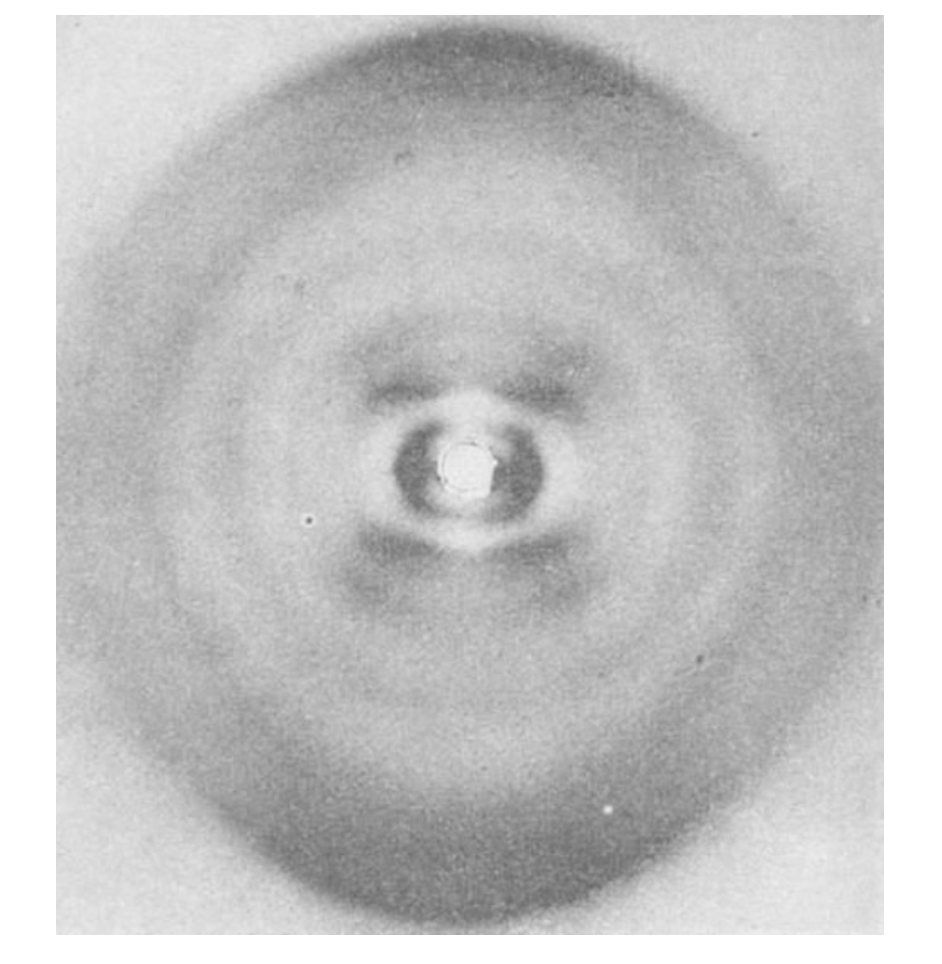 图 1. 来自大肠杆菌(B. coli)的脱氧戊糖核酸纤维图谱。纤维轴呈垂直方向。
图 1. 来自大肠杆菌(B. coli)的脱氧戊糖核酸纤维图谱。纤维轴呈垂直方向。通过每个贝塞尔函数的内侧极大值和原点,可以近似画出一条直线 。该直线与赤道的夹角大致等于螺旋元素与螺旋轴之间的夹角 。如果一个单元沿螺旋重复 $n$ 次,则在第 $n$ 条层线上会出现一个子午反射($J_0^2$)。螺旋构型在这一基频上产生边带,其效果是在第 $n$ 条层线上围绕新原点(对应图 2 中的 C)重现原点周围的强度分布 。
我们现在简要地分析核苷酸形状和大小对衍射图谱的一些物理影响 :
- 如果核苷酸由一个关于平行于螺旋轴的轴对称单元组成,整个衍射图谱都会受到核苷酸形状因子的修正 。
- 如果核苷酸由垂直于螺旋轴半径上的一系列点组成,则穿过每个点的不同直径螺旋所散射的辐射相位是相同的 。相应贝塞尔函数的求和会增强内侧极大值,而通常会因相位差而抵消所有其他极大值 。
- 如果核苷酸在垂直于螺旋轴的平面内呈圆弧状延伸,由于来自核苷酸上各点螺旋的辐射相位差,从原点发出的贝塞尔函数层线条纹强度会受到修正 。
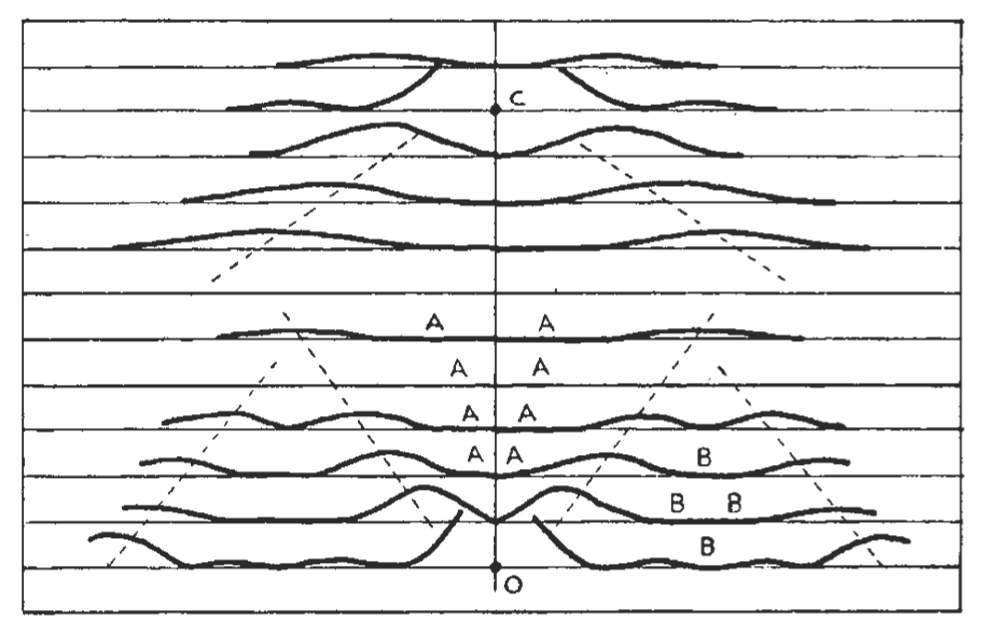 图 2. 对应于脱氧戊糖核酸结构的螺旋系统的衍射模式。图中在赤道线以及第一、第二、第三和第五层线上,围绕原点 O 绘制了贝塞尔函数(Bessel functions)的平方。计算假设一半的核苷酸质量分布在 20 Å 直径处,其余部分沿半径分布,且给定半径处的质量与该半径成正比。在第十层线的 C 点附近,针对 12 Å 的外径绘制了类似的函数。
图 2. 对应于脱氧戊糖核酸结构的螺旋系统的衍射模式。图中在赤道线以及第一、第二、第三和第五层线上,围绕原点 O 绘制了贝塞尔函数(Bessel functions)的平方。计算假设一半的核苷酸质量分布在 20 Å 直径处,其余部分沿半径分布,且给定半径处的质量与该半径成正比。在第十层线的 C 点附近,针对 12 Å 的外径绘制了类似的函数。必须首先确定该结构主要由一个螺旋组成(层线强度分布对应 $J_1, J_2, J_3…$),还是由两个相似且同轴、尺寸为上述两倍且沿轴向错开半个螺距的螺旋组成(给出 $J_2, J_4…$),或是三个螺旋等 。通过检查层线条纹的宽度,发现强度更接近于 $J_1^2, J_2^2, J_3^2$ 。因此,主螺旋的螺距约为 34 Å,根据螺旋角度计算,其直径约为 20 Å 。
为了在纤维中获得合理数量的核苷酸,需要两到三个相互缠绕的同轴螺旋,每个螺旋的一圈上有 10 个核苷酸 。子午线上或附近的反射缺失(图 2 中的空白区域 AAA)是螺旋结构的直接后果 。照片中赤道及其附近也存在一个相对空旷的区域(图 2 中的区域 BBB),这种次级贝塞尔极大值的缺失可以通过核苷酸形状的径向分布来产生 。
为了使层线条纹足够窄,必须将大部分核苷酸质量放置在约 20 Å 直径处 。图 2 绘制了假设一半质量在 20 Å 直径,其余沿半径分布的情况 。在第零层线上似乎有显著的 $J_{10}$,这表明在垂直于纤维轴的投影平面上,核苷酸的外部相对集中,在 20 Å 直径的圆周上产生间距约 6 Å 的高密度区域 。在第五层线上,两个 $J_5$ 函数重叠并产生强烈反射 。这暗示氮基在螺旋系统的中心区域像“一堆硬币”一样排列 。
证明上述螺旋结构确实存在于完整的生物系统中的证据简述如下:
- 精子头:晶体状精子头的 X 射线光谱强度由螺旋形式函数决定 。离心的鳟鱼精液与之前使用的干、湿精子头显示出相同的模式
- 噬菌体:在带有云母窗口的密封池中拍摄的 $T_2$ 噬菌体湿沉淀照片显示的衍射图谱,包含了准晶钠核酸盐的主要特征 。这证实了目前关于噬菌体结构的观点 。
- 转化因子:在 60% 湿度下干燥的活性脱氧戊糖核酸具有与某些胸腺核酸钠样本相同的晶体结构 。
虽然上述讨论并非完全没有疑义,但总体而言,实验数据与沃森(Watson)和克里克(Crick)描述的模型类型之间存在合理的一致性 。值得注意的是,如果每个直径 20 Å、螺距 34 Å 的螺旋上有 10 个磷酸基团,那么磷酸酯骨架链几乎处于完全伸展的状态 。因此,当钠核酸盐纤维被拉伸时,螺旋显然像处于张力下的螺旋弹簧一样被拉长 。
M. H. F. Wilkins
医学研究委员会生物物理研究单位
A. R. Stokes
H. R. Wilson
伦敦国王学院(King’s College, London),惠斯通物理实验室
1953 年 4 月 2 日
第三篇:精确实验数据
胸腺嘧啶核酸钠的分子构型(第 740–741 页)
作者:Rosalind E. Franklin & R. G. Gosling
胸腺嘧啶核酸钠纤维给出两种不同类型的 X 射线图谱 。第一种对应于一种晶体形态,即“结构 A”,在约 75% 的相对湿度下获得;关于此结构的详细研究已在别处阐述 。在更高的湿度下,会出现一种不同的结构,即“结构 B”,它显示出较低的有序度,并在较宽的环境湿度范围内保持稳定 。
A 到 B 的转变是可逆的 。经历这种可逆变化的结构 B 纤维,其含水量可从干重的 40%–50% 变化到百分之几百 。此外,某些纤维从未显示出结构 A,在这些纤维中,即使在更低的含水量下也能获得结构 B 。
结构 B 的 X 射线图谱(见照片)以显著的方式展示了螺旋结构的特征,这些特征最初由本实验室的斯托克斯(Stokes,未发表)以及克里克(Crick)、科克伦(Cochran)和范德(Vand)推导出 。斯托克斯和威尔金斯(Wilkins)是基于对核酸纤维的直接研究首先提出这种螺旋结构的,尽管此前富伯格(Furberg,1949年伦敦论文)基于对核苷和核苷酸的 X 射线研究也曾建议过螺旋结构 。
虽然目前 X 射线证据尚不能直接证明该结构是螺旋状的,但下文讨论的其他因素使螺旋结构的存在具有高度可能性 。当胸腺嘧啶核酸钠纤维吸收超过其重量约 40% 的水时,结构 B 便从晶体结构 A 衍生而来 。这一转变伴随着纤维长度增加约 30%,以及分子发生实质性的重排 。因此,可以合理地假设在结构 B 中,胸腺嘧啶核酸钠的结构单元(分子或分子组)相对不受相邻分子的影响,每个单元都被一层水鞘所屏蔽 。随后每个单元都能独立于其邻居采取其最低能量构型,鉴于所涉及的长链分子性质,其总体形式极有可能是螺旋状的 。
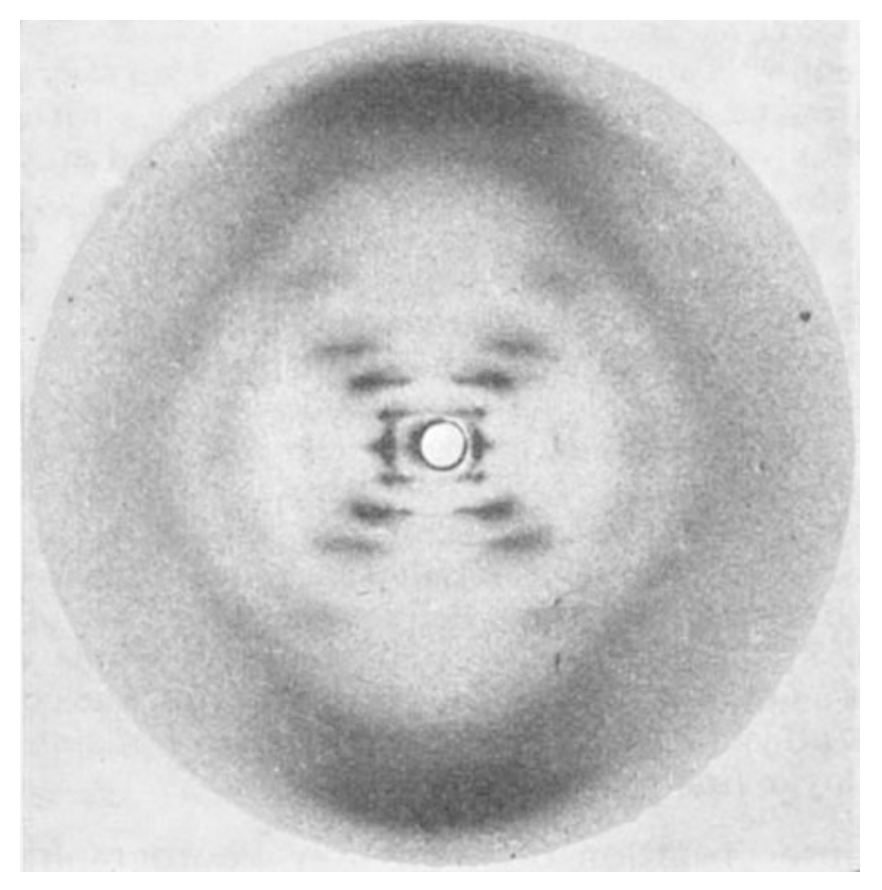 图 1. 胸腺嘧啶核酸钠的纤维 X 射线衍射图谱,结构 B(这就是科学史上著名的 Photo 51,它直接证明了 DNA 的螺旋结构)
图 1. 胸腺嘧啶核酸钠的纤维 X 射线衍射图谱,结构 B(这就是科学史上著名的 Photo 51,它直接证明了 DNA 的螺旋结构)如果我们采用螺旋结构的假设,立刻就能从结构 B 的 X 射线图中对螺旋的性质和尺寸做出某些推论 。在第一、第二、第三和第五层线上,最内侧的极大值点近似位于从原点辐射出的直线上 。对于平滑的单股螺旋,第 $n$ 层线上的结构因子由下式给出:
$$F_n = J_n(2\pi rR) \exp in(\psi + \frac{1}{2}\pi)$$
其中 $J_n(u)$ 是 $u$ 的 $n$ 阶贝塞尔函数,$r$ 是螺旋半径,$R$ 和 $\psi$ 是倒易空间中的径向和方位坐标 ;该表达式产生了一个观察到的近似线性的强度极大值阵列,对应于函数 $J_1, J_2, J_3$ 等的首个极大值 。
如果考虑的是沿螺旋等距分布的一系列残基,而非平滑螺旋,则由克里克、科克伦和范德处理的一般情况下的变换会更复杂 。但如果每圈有整数 $m$ 个残基,其变换形式与平滑螺旋相同,只需在高度 $mc^*, 2mc^*$ 等处重复相同的模式($c^*$ 为纤维轴周期) 。在目前的情况下,纤维轴周期为 34 A 。3.4 A 处的极强反射位于第十层线上 。此外,从 3.4 A 反射处辐射出的极大值线在第五层及更低层线上可见,且在第五层线上与原点系列具有重合的 $J_5$ 极大值 。这有力地表明螺旋每圈恰好有 10 个残基 。
若是如此,通过测量 $R_n$(第 $n$ 层线上首个极大值的位置,$n \le 5$),即可获得螺旋半径 $r$ 。在当前实例中,对 $R_1, R_2, R_3$ 和 $R_5$ 的测量均得出 $r$ 约为 10 A 的值 。由于这一线性极大值阵列是 X 射线图中最重要的特征之一,我们必须断定分子的一个晶体学上的重要部分位于该直径的螺旋上 。这只能是磷酸基团或磷原子 。
如果 10 个磷原子位于半径 10 A、螺距 34 A 的螺旋的一圈上,分子中相邻磷原子间的距离为 7.1 A 。这对应于完全伸展分子中的 P…P 距离,因此进一步表明磷酸基团位于结构单元的外侧 。
因此,我们的结论不同于鲍林(Pauling)和科里(Corey),他们曾为核酸提出一种磷酸基团构成致密核心的螺旋结构 。
现在我们必须简要考虑赤道反射 。对于单螺旋,赤道极大值系列应对应于 $J_0(2\pi rR)$ 的极大值 。然而,照片上的极大值并不符合基于上述推导出的 $r$ 值的函数 。在约 24 A 处有一个极强反射,随后仅在 9.0 A 处有一个微弱尖锐反射,以及在 5.5 A 和 4.0 A 附近有两个弥散带 。然而,这种不符是在预料之中的,因为我们知道迄今为止考虑的螺旋只能是不同半径的一系列同轴螺旋中最重要的一员 ;分子的非磷酸部分将位于内部同轴螺旋上,可以证明,虽然这些部分不会显著影响层线上的最内侧极大值,但它们可能会破坏或移动赤道极大值以及其他层线上的外侧极大值 。
因此,如果结构是螺旋状的,我们发现磷酸基团或磷原子位于直径约 20 A 的螺旋上,糖基和碱基基团也相应地向内指向螺旋轴 。然而,密度考量显示,高度 34 A、直径 20 A 的圆柱体重复单元包含的核苷酸必须远多于 10 个 。由于结构 B 常存在于低含水量的纤维中,螺旋单元的密度似乎与干胸腺嘧啶核酸钠的密度(1.63 g/cm³)差异不大,高含水量纤维中的水分位于结构单元外部 。在此基础上,我们发现半径 10 A、高度 34 A 的圆柱体将包含 32 个核苷酸 。
因此,仅凭密度测量很难确定一个重复单元是在两个还是三个同轴分子上各包含 10 个核苷酸 。然而,另外两个论据使只有两个同轴分子的可能性极高 。首先,使用叠加法对结构 A 的帕特森函数进行研究表明,该结构的基元晶胞中只有两条链通过 。由于 $A \rightleftharpoons B$ 转变易于逆转,分子在结构 B 中以三条一组排列的可能性极小 。其次,从结构 B 的 X 射线图测量可以证明,无论每个单元的链数是二还是三,这些链在纤维轴方向上并非等距分布 。例如,等距的三条链意味着第 $n$ 层线取决于 $J_{3n}$,这将导致直径约 60 A 的螺旋,这比结构 A 的晶胞大得多,且相对于核苷酸尺寸而言大得离谱 。因此,很可能只有两个同轴分子,且它们在纤维轴方向上不等距分布 。
综上所述,虽然我们不打算对结构 B 的纤维图谱提供完整解释,但可以陈述以下结论 :该结构可能是螺旋状的 。磷酸基团位于结构单元的外侧,处于直径约 20 A 的螺旋上 。结构单元可能由两个同轴分子组成,它们在纤维轴方向上不等距分布,其相互位移解释了层线上观察到的最内侧极大值强度的变化;如果一个分子相对于另一个分子位移约纤维轴周期的 3/8,这将解释第四层线极大值的缺失和第六层线的微弱 。
因此,我们的总体思路与前一篇文章中沃森(Watson)和克里克(Crick)提出的模型并无矛盾 。磷酸基团位于结构单元外侧的结论此前已通过完全不同的推理得出 。第一条论据源于古兰德(Gulland)及其合作者的工作,他们表明即使在水溶液中,碱基的 -CO 和 -NH₂ 基团也是不可接近且无法滴定的,而磷酸基团则是完全可接近的 。第二条论据基于我们自己的观察,即结构 A 和 B 中的结构单元如何被过量水分逐步分离,这一过程是连续的,最终导致凝胶甚至溶液的形成 。
我们感谢 J. T. Randall 教授的支持,并感谢 F. H. C. Crick 博士、A. R. Stokes 博士和 M. H. F. Wilkins 博士的讨论 。
罗莎琳德·E·富兰克林
R·G·葛斯林
伦敦国王学院,惠斯通物理实验室